光合成においてどのような物理化学的反応が進行するかは、
別の所で解説していますので、ここでは、エントロピーの問題に限って説明します。
途中で関与する化学物質を無視して変化する部分だけを抜き出すと、光合成の化学反応は、次の簡単な化学式で表すことができます。
6CO
2 + 6H
2O → C
6H
12O
6 + 6O
2
はじめは大気中に二酸化炭素として広がっていた炭素原子が、光合成によって炭水化物として固定されるわけですから、物質だけを見る限り、エントロピーは減少していると考えられます。各物質の熱力学的なエントロピーを使って計算すると、この化学反応を通じて、炭水化物1モル当たり 70R (R:気体定数)のエントロピー減少があることがわかります。
物質系におけるエントロピーの減少を補償して、全体としてエントロピーを増大させているのが、外界との間で行われる吸光・放熱の過程です。太陽からの放射は、表面温度6000K の(宇宙空間に比べて)小さな領域から、3K の背景放射を持つ広大な宇宙空間へと光エネルギーが流れ出す非平衡過程であり、莫大なエントロピー生成を伴っています。このエネルギー流に巻き込まれる形で生じるサブプロセスの中には、全体系でのエントロピーを減らさずに、部分系でのエントロピーを減少させているものもあります。光合成は、そうしたサブプロセスの1つなのです。
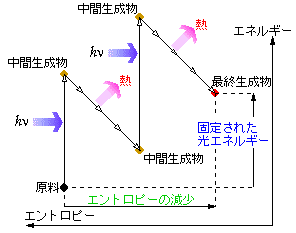
光合成は、2段階の吸光・放熱過程から成っており、その間の物質側におけるエントロピー増減は、概略的には右図のようになっています。物質と電磁場が平衡状態になっていれば、逆過程も同じように生じるので原料から最終生成物への一方向的な変化はありませんが、短波長領域で著しい非平衡状態になっているため、非可逆的な合成が自発的に進行していくことになります。
なお、上では「放熱」という言い方をしましたが、現実の反応では、根から吸い上げられた液体の水が気孔から蒸散することによって、葉の温度を上げずに熱を奪っていきます。水分子1モルの蒸発によるエントロピー増大は 18R なので、炭水化物1モル合成するために減少する 70R のエントロピーを補償するには少なくとも 4モルの水の蒸発が必要ですが、実際には、光合成反応と関連して数百モルの水分子が蒸発しているようです。したがって、エントロピーのことまで考えた光合成の反応式は、次のように書くべきでしょう。
6CO
2(気体) + 6H
2O(液体) + (数百)H
2O(液体)+ 短波長光
→ C
6H
12O
6 + 6O
2(気体) + (数百)H
2O(気体)
(この回答の執筆には、勝木渥著『環境の基礎理論』(海鳴社)を参考にしました)
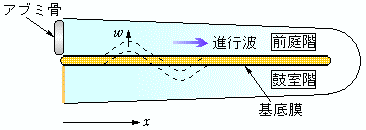
内耳にある蝸牛は、カタツムリの殻のようにらせん形をしており、横断面を見ると、基底膜によって前庭階と鼓室階の2つの部分
(*)に分かれ、それぞれリンパ液で満たされています。中耳のアブミ骨から蝸牛に伝えられた音波の振動は、リンパ液に圧力差を生じさせ、基底膜の振動を引き起こします。このとき、周波数に応じて基底膜の特定の場所(分節)が最も激しく振動するため、そこにある神経線維が刺激されてインパルスを発射します。振動する場所は、高周波では蝸牛の入り口近く(1000Hzではアブミ骨から20mm付近)、低周波では奥の方(100Hzで30mm付近)になります。これが、「ある高さの音が聞こえる」という認知の元になるわけです。
(*)正確には、中央階を加えた3つの部分に分かれていますが、ここでは話を簡単にしています。
基底膜が振動するプロセスはかなり複雑で、膜自体が能動的に振動を増幅するような生理的なメカニズムも関与しています。しかし、周波数と振動する場所の対応関係を明らかにするだけならば、きわめて単純な物理学的モデルで充分です。このモデルでは、蝸牛はまっすぐに伸びているとされ、振動に関する力学的な量は、中心軸の位置座標xだけの関数として表されます。リンパ液を非圧縮性の完全流体と仮定すると、蝸牛内部を伝わる波は、水の表面波と同じように扱うことができます。ただし、水の表面波の場合、変位を元に戻そうとする作用は重力が与えますが、蝸牛内部の進行波では、基底膜が基準位置からずれることによって生じる弾性力(=変位×弾性定数)が復元力となります。さらに、線形近似を採用すれば、入射する音波の角振動数(周波数×2π)をωとすると、すべての力学変数の時間依存性は、 exp(iωt) で与えられます。したがって、方程式に現れる時間微分は iω で、時間積分は(積分定数を別にして) 1/iω で置き換えることができます。
波の解析を行うのに必要なのは、リンパ液の連続の式と(中心軸方向の)運動方程式、それに音響インピーダンスの式の3つですが、周波数と振動が大きくなる場所の対応関係は、3番目の式だけで求められます。音響インピーダンスζ(x) は、前庭階と鼓室階の圧力差 p(x) と基底膜の垂直方向の速度 w(x) の比として定義され、波動の伝播に対する抵抗と考えることができます(いずれも時間依存性 exp(iωt) を除いた複素数として定義されることに注意)。
ζ = -p/w
ここで考えているモデルの場合、音響インピーダンスは3つの項に分解されます。
ζ = iωL(x) + 1/iωC(x) + r(x)
第1項はリンパ液や基底膜の慣性による抵抗を表しており、単位長さあたりの質量を L(x) としたときの垂直方向の慣性力 Ldw/dt の時間微分を iω で置き換えれば得られます。第2項は、基底膜の変位によって生じる弾性力fの作用を、圧力差に加えて考慮しなければならないことを表しています。基底膜の弾性定数を 1/C(x) とすると、
f = -∫wdt /C
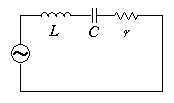
となり、時間積分を 1/iω で置き換えれば上の式が得られます。第3項は、同位相で作用する抵抗をまとめたものです。
この音響インピーダンスは、交流電源に、インダクタンスL のコイル、容量C のコンデンサー、抵抗r の抵抗器を直列に接続した回路のインピーダンスと同じ形をしています。等価回路と同様にインピーダンスの虚部がゼロになる角振動数
ω = 1/(LC)
1/2
で、共振を起こします。
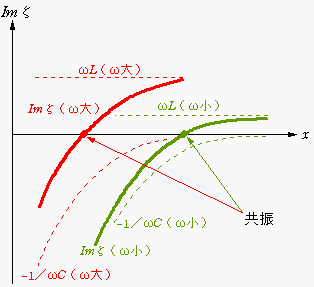
実際の蝸牛において、質量 L(x) は場所によってほとんど変わらないのに対して、弾性定数 1/C(x) は、蝸牛の入り口付近で大きく、奥になるに従って急速に小さくなります。したがって、音響インピーダンスの虚部 Imζ は下の図のようになり、ωの振動が入力されたときに共振を起こす位置x は、ωが大きいほど入り口寄りになります。これが、高周波数の音波が蝸牛の入り口近くで感知される理由です。直観的に言えば、蝸牛の奥の方は弾性定数が小さく慣性の効果が相対的に強く効くため、物体を速く振動させようとする高周波ほど進入しにくくなるということです。
蝸牛の内部で波がどのように伝わっていくかを調べるには、音響インピーダンスの式の他に、リンパ液の連続の式と運動方程式を連立させる必要があります。この連立方程式を数値的に解くと、進行波の振幅がしだいに大きくなり、ある地点で最大値を取った後に急激に減衰する解が得られます(詳しくは、E.de Boer, Physics Reports 62(1980)87などを参照)。こうした進行波の振舞いは、BeKesy が1960年に死体の内耳を使って行った観察の結果と定性的に一致しており、上の単純なモデルが現実的なものであることを示しています。
通常、双極子と言えば、電気双極子または磁気双極子のどちらかを指します。
有限の領域内部に電荷が集まっているとき、そこから充分に離れた地点での電位(静電ポテンシャル)φは、全電荷Qがゼロでなければ、近似的にクーロンの法則に従って、電荷のある領域までの距離Rに反比例して減少します。
φ ≒ Q/R
しかし、Q=0 の場合は、φは近似的にR
2に逆比例し、次のように表されることが知られています。
φ ≒
d・
n/R
2
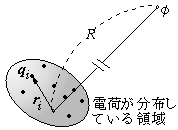
ただし、
nは電荷の存在する領域から電位を測る地点までの方向ベクトルです(上の式は、φをRの逆数で展開したときの2次の項と見なしても良い)。このとき、
dが(電気的な)双極子モーメントで、大きさq
iの点電荷が位置
riに存在している場合は、
d = Σq
iri
で与えられます。Q=Σq
i=0の場合、
dは位置ベクトルの原点の選び方に依存しません。特に、+qと-qという2つの点電荷だけが
rだけ離れて存在するときには、
d = q
r
という簡単な関係が成立します。
磁気の場合、N(S)極だけの単極子(磁荷)は存在せず、物質から充分に離れた地点での磁場は、(磁気的な)双極子モーメント
mによって支配されます。
mが作るベクトル・ポテンシャル
Aは、
A =
m×
n/R
2
(×は外積を表す)となります。磁気双極子の起源は、電子のスピンや原子内部の電子の運動、あるいは、物質内部に流れるループ状の電流です。
二重らせんを形成するDNAの構造には、いくつかの種類(A型・Z型など)がありますが、水中でエネルギー的に最も安定なのは、B型と呼ばれる右巻きらせんで、線状DNAの場合、塩基10.4個ごとにらせんが1巻きします。ところが、細菌やミトコンドリアのDNAは、線状DNAの両端がつながった閉じた環の形をしています。こうした環状DNAでは、例えば、「塩基数が1040個で、らせんがちょうど100回巻いている」ということは稀で、通常は、塩基数に対して、らせんの巻き数が足りない(1040個の塩基に対して95回しか巻いていないような)状態になっています。環状DNAのらせんの巻き数は、DNAをいったん切断しない限りは変えられないので、二重らせんの中心軸自体がねじれることによって、巻き数が足りないことによって生じた歪みを少しでも解消しようとします。こうしてできるのが負の超らせん構造で、生きた細胞中の環状DNAは、全てこの構造を取っています。
負の超らせん構造を持つDNAは、“弛緩した”B型DNAにはないいくつかの特長を持っています。第1に、サイズがきわめてコンパクトになり、全遺伝情報がコードされたDNA全体を細胞内部にすっぽりと納めることができます。第2に、負の超らせん構造は2本鎖の分離を容易にするため、DNAの複製が促進されるという性質があります。こうした利点を最大限に生かそうとして、生物は、酵素を使って超らせん構造の形成や解消を能動的に制御しています。こうした酵素はトポイソメラーゼと呼ばれ、あらゆる生物に備わっています。